摘要 摘 要: 土壤微生物对土壤变化的反应极为敏感,秸秆还田对土壤微生物产生的影响对维持稻田系统氮素循环极其重要。秸秆还田通过对土壤微生态环境的改变影响氮素在土壤中的生物化
摘 要: 土壤微生物对土壤变化的反应极为敏感,秸秆还田对土壤微生物产生的影响对维持稻田系统氮素循环极其重要。秸秆还田通过对土壤微生态环境的改变影响氮素在土壤中的生物化学转化过程,进而影响水稻生产的氮素管理策略。在查阅近年来发表的相关研究论文基础上,对秸秆还田下水稻土壤氮素转化过程中相关功能微生物的作用以及关键功能基因驱动机制等研究进行综述,以期厘清秸秆还田后稻田生态系统中微生物介导的氮素循环转化机制,为稻田科学施用氮肥提供理论依据。
关键词: 秸秆还田; 稻田; 氮循环; 土壤微生物; 功能基因
农作物秸秆富含有机物质和氮、磷、钾等营养元素。秸秆还田既可为作物提供所需养分,又能为土壤微生物提供丰富碳源,刺激微生物活性,进而促进氮循环和矿化,提高氮肥利用率,改善土壤培肥效果[1 - 2],因此,秸秆还田已成为农业可持续发展的重要措施之一。秸秆还田形式多样,主要有粉碎后直接翻压还田、覆盖还田、堆沤还田和腐熟剂处理还田等,其中,秸秆粉碎后直接翻压还田是当前秸秆肥料化利用的主要方式[3]。
稻田秸秆还田后会对土壤氮素的转化过程产生深刻影响,还田方式、还田年数、还田量,以及无机肥料配施方式等都是重要的影响因子。秸秆还田通过土壤氮素转化过程影响水稻的生长和产量。不同施肥处理对土壤铵态氮的影响不大,但施用氮肥会显著增加土壤中硝态氮的含量[4 - 5]。这可能是因为秸秆还田增加了土壤中的有机质含量,阻止了 NH + 4 的晶格固定,促进了氮的硝化[6]。Takahashi 等[7] 研究表明,短期内秸秆对氮素和土壤无机氮的贡献不足,秸秆施用对作物氮素吸收和氮素矿化的影响很小。同时,苯酚( 源自部分降解的木质素和一些真菌色素) 可以与氨基酸或氨反应形成含氮的腐殖质聚合物,阻碍土壤微生物的降解,导致植物处于快速生长阶段且具有相对高的氮需求时氮供应不足。秸秆还田 4 ~ 6 a 后则会形成一个不稳定的有效氮库,降低水稻对氮肥的依赖性[8],当还田年限达到 10 a 以上,土壤养分趋于稳定[9]。稻田系统有淹水和干湿交替的过程,相应的氮素转化呈现诸多特点[10],主要变化有: 土壤铵态氮在水稻苗期和分蘖期含量较高,之后呈下降趋势; 硝态氮在齐穗期浅层土壤和苗期深层土壤中含量较稳定,其余生育期波动较大[11]; 淹水期土壤铵态氮含量较高[12]; 苗期由于根部细胞色素氧化酶占优势且根内缺少硝酸还原酶,水稻吸收铵态氮较多,但随着生育进程,水稻体内硝酸还原酶经诱导逐渐 生 成 并 占 据 优 势,对硝态氮的吸收增加[4]; 水稻收获后 0—60 cm 土层铵态氮含量明显高于硝态氮,且铵态氮随土层深度的增加而递增。
上述这些土壤中的氮素转化大多在微生物驱动下进行,因此越来越多的研究者开始关注微生物介导下的水稻土中各种形态氮素的转化过程及其相互影响机制。本文就近年来国内外秸秆还田模式下微生物介导的稻田氮素转化的微生物群落及功能基因研究进行梳理,以期为秸秆还田提高氮肥利用率提供更多理论指导。
1 秸秆还田对稻田微生物群落的影响
研究表明,秸秆还田配施化肥能显著提高土壤微生物数量,尤其是氨氧化细菌、固氮菌、硝化细菌、反硝化细菌和纤维分解菌等氮素转化功能菌的数量,进而对土壤氮素循环产生重要影响。但也有研究表明,秸秆还田会减少土壤有机质积累和微生物多样性[3],对土壤微生物群落组成和功能产生强烈影响[13]。随着秸秆不断分解,土壤微生物可利用的营养物质数量减少,会进一步抑制土壤微生物活性[14]。土壤微生物的数量及活性主要取决于土地利用类型、pH 值及有机质含量,当这些因素不同,且秸秆还田量和年数不同时,其对稻田微生物群落的影响也不尽相同[15]。
1. 1 pH 值在 pH 值 5. 77 的成都平原区,秸秆还田翻耕处理的土壤氨氧化细菌、硝化细菌和固氮菌分别较对照增加了 1. 9 倍、1. 8 倍和 22. 7 倍,免耕处理的固氮菌增加了 25. 0 倍,但氨氧化细菌和硝化细菌的数量略有降低; 在 pH 值 6. 38 的川中丘陵区,翻耕处理的氨氧化细菌、硝化细菌和固氮菌数量分别较对照增加了 25. 0 倍、2. 6 倍和 7. 5 倍,免耕处理的氨氧化细菌、硝化细菌和固氮菌分别较对照增加了 10. 4 倍、1. 7 倍和 1. 5 倍[16]。
1. 2 有机质含量川中丘陵区秸秆还田处理较对照减少了土壤反硝化细菌数量,但在有机质含量较高的平原区 无 此 明 显 规 律。这可能与有机碳 含量有关[16]。土壤有机质含量的提高会增加土壤微生物碳含量及微生物数量,促进活性酶的产生[17]。
1. 3 土地利用类型在长期连作土壤中,秸秆深埋较秸秆覆盖更能增加土壤细菌、放线菌、氨氧化细菌、好气性固氮菌、硝化细菌的数量,而在轮作土壤中,秸秆还田方式对根际土壤真菌和主要微生物生理类群的影响不大。长期连作后,土壤微生物单一,多样性差,细菌生理类群数量相对较少。秸秆深埋后,其腐解速率大于秸秆覆盖还田处理,能改善耕作的犁底层,保蓄水分,有效提高土壤有益微生物的数量和酶活性[18]。
1. 4 秸秆还田量通常高秸秆还田率易改变微生物群落结构,提高大部分水解酶的活性,而低秸秆还田率对其影响较小[19]。秸秆还田对微生物的影响随着还田量的增加先增加后减小,翻耕条件下以 2 /3 还田量处理的土壤微生物数量和活性较高,免耕条件下 1 /3 还田量处理的提高作用显著[20]。
1. 5 秸秆还田年限短期秸秆还田对作物氮和土壤无机氮的贡献率较低,对淹水条件下土壤氮素矿化和作物氮素吸收影响不大; 长期秸秆还田可以改善土壤肥力,促进作物生长,加快氮素吸收[21]。因此,试验年数不足可能会影响相关结果的准确性。
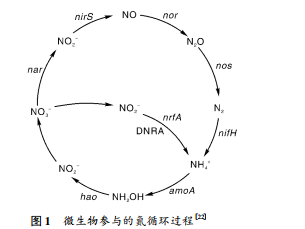
2 秸秆还田对稻田氮素转化功能基因的影响土壤的氮素循环主要包括固氮作用、硝化作用、反硝化作用和铵还原过程,主要由固氮微生物、硝化功能菌、反硝化功能菌和硝酸盐异化还原成铵过程( DNRA) 功能微生物驱动[22],如图 1 所示。目前,广泛使用分子标记,如 nifH、amoA、 nirK、nirS 和 nrfA 基因来研究土壤氮循环微生物。氮循环过程对应的基因包括: 氮固定的 nifH 基因,硝化作用的 amoA 基因,反硝化作用的 nirK 和 nirS 基因,以及 DNRA 的 nrfA 基因。它们的拷贝数通常被用来指示氮循环微生物丰度[23]。
2. 1 固氮微生物
图 1 微生物参与的氮循环过程[22] Fig. 1 Nitrogen cycling process by microorganism[22]固氮作用通常指将气态 N2 转化为 NH + 4 的过程。固氮微生物是稻田中重要的微生物功能菌群之一,通常存在于水稻根际生态系统中,对水稻氮素营养调节起着重要作用。目前发现的固氮微生物都属于原核生物界,分 59 个属: 好氧性自生固氮菌,包括固氮菌属( Azotobacter) 、固氮单胞菌属( Azotomonas) 等; 微好氧性自生固氮菌,包括德克斯氏菌属( Dexia) 、甲基单胞菌属( Methylomonas) 等; 兼性厌氧性自生固氮菌,包括芽孢杆菌属( Bacillus) 等; 专性厌氧性自生固氮菌,包括梭菌属( Clostridium) 、脱硫肠状菌属( Desulfotomaculum) 等; 好氧性联合固氮菌,包括拜叶林克氏菌属( Beijerinckia) 等; 微好氧性联合固氮菌,包括 粪 产 碱 菌 属 ( Alcaligenes ) 、节 杆 菌 属 ( Arthrobacter) 等; 兼性厌氧性联合固氮菌,包括肠杆菌属( Enterobacter) 、克雷伯氏菌属( Klebsiella) ; 光能自养型固氮菌,包括红细菌属( Rhodobacter) 、红螺菌属( Rhodospirillum) 等[24]。根据与其他生物的关系,这些微生物可分为自生固氮微生物、联合固氮微生物和共生固氮微生物。这些固氮微生物体内存在一种具有催化功能的固氮酶,其中,微生物固氮酶复合物包括 2 个保守蛋白,即由 nifD 和 nifK 基因编码的钼铁蛋白和由 nifH 基因编码的铁蛋白。nifH 基因负责编码固氮酶组分铁蛋白亚基,为固氮微生物所共有,具有高度保守性[25],是进化过程中最古老的功能基因之一,也是研究固氮微生物群落结构最好的标记基因[26]。
土壤中固氮基因 nifH 的多样性受到土壤耕作制度、植被类型、土壤理化性质等诸多因素的影响,只有综合考虑各个因素的效应才能提高土壤的固氮效率,进而提高土壤中氮肥的生物有效性。刘骁蒨等[27]研究表明,秸秆覆盖配施足够的氮磷钾肥能够维持土壤充足的营养物质,防止植物与微生物争氮,有利于固氮细菌的生长繁殖,且秸秆覆盖处理相对于无秸秆处理,固氮菌多样性指数及其丰度显著增加。杨滨娟等[28]研究发现,秸秆还田配施化肥能够增加好气性自生固氮菌的数量,而普通区组间差异不显著。秸秆还田后能够通过矿化和降解作用释放氮以补偿植物和微生物的可用氮库,固氮微生物可以直接从秸秆还田处理的土壤中吸收氮,不需要固氮消耗能量,因此秸秆还田可能会导致 nifH 基因表达水平和固氮活性下降。然而,有研究表明,秸秆还田处理的 nifH 基因拷贝数仍高于无秸秆还田处理[29 - 30]。这说明秸秆还田可以提高土壤自身固氮能力,对土壤肥力的提高产生持久影响。总体来看,这些研究主要集中在固氮菌数量和多样性的分析上,缺乏对秸秆不同还田方式和化肥配施下固氮基因响应机理和驱动因子的研究,因此很难明确何种秸秆还田方式能有效、持久地提高稻田固氮能力。
2. 2 硝化功能菌
硝化作用包括 2 个过程: 氨在氨氧化细菌 ( ammonia-oxidizing bacteria,AOB) 和氨氧化古菌 ( ammonia-oxidizing archaea,AOA) 的作用下被氧化为亚硝,然后亚硝在亚硝酸氧化细菌( nitriteoxidizing bacteria,NOB) 作用下被氧化为硝酸盐[31]。氨的氧化主要靠氨氧化细菌来完成,部分古细菌同样参与这一重要的生化反应过程。大量研究表明,氨氧化细菌主要集中于亚硝化单胞菌属( Nitrosomonas) 、亚 硝 化 弧 菌 属 ( Nitrosovibrio) 、亚硝化叶菌属( Nitrosolobus) 、亚硝化螺菌属( Nitrosospira) 和亚硝化球菌属( Nitrosococcus) 等 5 个属[32]。执行硝化作用第二步的亚硝酸氧化菌主要有硝化杆菌( Nitrobacter) 、硝化刺菌( Nitrospina) 、硝化球菌( Nitrococcus) 和硝化螺菌( Nitrospira) 。克隆文库分析显示,硝化螺菌在水稻土中丰度最高[33]。现在,有一种新的氨氧化过程———厌氧氨氧化在水稻土中被发现[34]。目前已知的厌氧氨氧化菌 Candidatus brocadia、Candidatus kuenenia、Candidatus scalindua、Candidatus anammoxoglobus、Candidatus jettenia 全 部 归 属 于 Brocadiales 目。
细菌 和 古 细 菌 均 具 有 编 码氨单加氧酶 ( AMO) 的 amoA 功能基因,其具有一定的序列保守性,通常用来标记硝化过程。编码经胺氧化酶 ( HAO) 的基因 hao,也是研究氨氧化阶段的标记基因。氨氧化细菌属于化能自养型微生物,广泛分布于湖泊、底泥、土壤和海洋环境中。水稻土常处于淹水状态,但水稻土表层含氧量较高,仍可进行硝化作用[35]。长期秸秆还田能提高水稻土中可利用碳含量,促进异养细菌生长,对氨的竞争利用使优势种群突出,非优势种群的发展被抑制,从而降低水稻土中 amoA 和 hao 基因群落的多样性,改变细菌群落组成,并导致氨氧化菌群的富集[36]。众多研究已经证实,有机碳是驱动 amoA 变化的重要因子。Jin 等[37]研究发现,有机肥或秸秆配施化肥会提高土壤中 amoA 和 nirK 的拷贝数,其中,amoA 与土壤有机碳、全氮有关,nirK 则与土壤有机碳、全氮、全磷相关。研究证明,秸秆还田配施化肥会通过有机碳的积累改变水稻土中硝化基因群落结构,提高硝化功能菌的丰度。因此,有研究者试图通过秸秆生物炭调节 amoA 基 因 来 控 制 水 稻 土 中 N2O 的 排放[38 - 39]。研究同时发现,稻田水分条件突变会影响 N2O 通量,影响氨氧化细菌和脱氮细菌群落生长的硝化细菌和脱氮细菌功能基因,随着含水量增加,功能基因丰度也会增加[40]。对于秸秆还田模式下的稻田,必须结合水分管理,厘清稻田中其他养分的影响,从而动态调控稻田中硝化基因的变化,平衡系统中的氮素有效转化,减少氮的排放与流失,提高氮素利用率。
2. 3 反硝化功能菌
反硝化是一个兼性厌氧的微生物过程,主要是将 NO - 3 或 NO - 2 还原为 N2O 或 N2,对环境有正负两方面的效应,既是温室气体 N2O 的主要来源,同时也是将活性氮转化为惰性氮( N2 ) 的唯一途径,通过去除过多的活性氮可减少硝态氮的淋溶损失,保持生态系统氮素平衡[41]。水稻根际反硝 化 细 菌 的 优 势属有假单胞菌属 ( Pseudomonas) 、芽孢杆菌属( Bacillus) 、微球菌属( Micrococcus) 、黄杆菌属( Flavobacterium) 、不动细菌属 ( Acinetobacter) 和肠细菌属 ( Enterobacter) 等,其中,以假单胞菌属最为常见,反硝化作用能力最强。根面以 Ps. aeroginosa 为主,根际则是 Acinetobacter sp. 、Ps. fluorescens 和 Ps. stutzeri 等较多[42]。
细菌、真菌和古细菌等种类繁多的微生物均参与 反 硝 化 过 程[43],且是在硝酸盐还原酶 ( nar) 、亚硝酸盐还原酶 ( nir) 、氧 化 氮 还 原 酶 ( nor) 和氧化亚氮还原酶( nos) 4 种酶的连续催化下完成的[32]。反硝化过程也可以看作是不同种类微生物共同作用的结果,因此对反硝化微生物的研 究 相 对 困 难[33]。目 前,亚 硝 酸 盐 还 原 酶 nirS、nirK 基因和氧化亚氮还原酶 nosZ 常被用作反硝化细菌的分子标记。反硝化细菌是一类异养型兼性厌氧细菌。在土壤中,反硝化作用主要受到盐度、pH、溶解性无机氮( DIN) 、溶解性有机碳( DOC) 、温度的影响[22]。水稻淹水期以厌氧环境为主,因此反硝化作用是稻田氮素转化的主要途径[34]。反硝化速率与反硝化微生物的丰度并不总是相关的。反硝化是一种机会性生长机制,可以依赖其他电子受体而活跃和生长。当前 DNA 数据只能证实微生物存在的信息,反映环境因素变化对微生物群落结构的影响,却没有关于代谢活性的数据[44]。蔚霞[41]研究发现,秸秆还田后土壤中反硝化速率降低,这可能与该实验没有配施氮肥有关,氮肥施用会促进土壤反硝化作用。王沛譞等[32]研究发现,转 cry1Ac/cpti 双价抗虫基因水稻的秸秆还田降解对土壤氮素转化关键微生物群落丰度与组成没有明显影响,不存在影响土壤氮素转化及 N2O 排放的可能,但由于土壤中产生 N2O 的途径多样,且有多种酶参与反应,故还需要进一步的研究来深入揭示转基因秸秆对稻田土壤 N2O 排放的影响及机制。Azziz 等[44]研究发现,nirS 型反硝化菌群落结构对环境因子比 nirK 型更敏感,有机质含量是影响 nirS 型反硝化菌群落结构的主要因素,而 nirK 基因不受土壤类型和水稻品种的影响,nirS 和 nirK 基因丰度会随着水稻生长周期的变化而变化。
近年来关于稻田反硝化基因的研究很多是围绕 N2O 排放展开的,重在探究水稻土中氮通量、N2O 浓度以及其中的微生物驱动机制[42,45]。 Cucu 等[46]研究发现,施用氮的气体损失( 主要为 N2 ) 在无秸秆添加时高达 60% ,而在有秸秆时仅为 20% ,说明连续秸秆还田会减少 N2O 排放。古菌 nosZ 基因对反硝化作用的高适应性可能会成为减少 N2O 排放的一个关键特征,同时在秸秆还田的情况下,细菌 nosZ 基因也会促进反硝化作用的完成。这些研究都表明,秸秆还田配施化肥会导致反硝化速率提高。反硝化底物包括氮素和有机质,秸秆进入土壤能显著提高土壤有机质储量,明显改善土壤有机质的活性和质量[47]。化肥和秸秆都可以作为氮源为微生物提供养分,但其中不同基因对环境因子的响应程度不一。要减少秸秆还田对环境造成的负面影响,深入细致探讨介导反硝化每一个步骤中的关键基因显得尤为重要。
2. 4 DNRA 功能菌
DNRA 是指厌氧条件下,NO - 3 还原为 NO - 2 和 NH + 4 的过程,且 NH + 4 为主导产物[48]。研究人员常用功能基因 nrfA 探究该过程,并研究 DNRA 微生物的多样性[24]。能够进行 DNRA 的细菌包括专性厌氧菌、兼性厌氧菌和需氧菌[49]。由于 Woods [50]发现 DNRA 可能发生在常见的土壤细菌如维氏梭菌( Clostridium welchii) 中,所以一些研究认为 DNRA 挑战了普遍认为反硝化基本上解释了厌氧土壤中所有 NO - 3 异 化 的观点[51 - 52]。
DNRA 是一个发酵过程,其中,NO - 3 还原与碳氧化相结合,因此在高活性碳和低 Eh 的环境中是有利的[53]。葡萄糖能显著提高 DNRA 速率[49],游离硫化物( H2 S、S2 - ) 可以作为 DNRA 的潜在电子供体,同时作为强还原剂会降低土壤 Eh,将 NO - 3 化学还原为 NH + 4 ; 但当硫化物与葡萄糖同时存在时,硫化物会显著提高未添加葡萄糖土壤中的 DNRA 速率,而对有葡萄糖添加的土壤无显著影响,说明葡萄糖会抑制由 S 驱动的 DNRA[54]。陈韬等[55]研究发现,当 Eh 在 - 225 ~- 340 mV 时,土壤氧化态降低,还原性增强,DNRA 作用生成的15 N-NH + 4 明显提高,反硝化作用减弱。杨雪琴等[22]研究表明,DNRA 与反硝化相比,在电子供体丰富、可利用 NO - 3 浓度低和温度高的环境中占主要优势。pH 对 DNRA 也有明显影响,pH 在 6. 2 ~ 8. 2 范围内的 DNRA 比率明显高于其他 pH 条件下[56 - 57]。此外,硝态氮含量和 C /N 也是影响 DNRA 的主要因素,C /N 越高对 DNRA 越有利[48]。
DNRA 不同于反硝化和厌氧氨氧化过程: 在河流沉积物硝酸盐异化过程中,DNRA 占比最小,反硝化、厌氧氨氧化和 DNRA 的占比分别为 50. 2% 、40. 5% 、9. 3%[48],而 Yin 等[49] 和 Lu 等[53]在水稻土中发现 DNRA 占比较高,分别为 15. 9% 和 3. 88% ~ 25. 4% 。因此,在特定条件下 DNRA 可能在稻田中发挥重要作用。Tatti 等[58]研究发现,一些细菌能够同时进行反硝化和 DNRA 作用,同时携带 nrfA 和 nirS /nirK 基因,且随着秸秆还田量的增加,DNRA 比例增加。秸秆还田通过提高土壤 DOC 含量,使 NO - 3 -N 的比例上升,进而提高 DNRA 速率[59]。Wang 等[60]研究也证明,秸秆还田为土壤提供了丰富的可利用碳, NO - 3 消耗从以固定化为主转移到以 DNRA 为主导。受茬管理导致的土壤环境差异也会影响水稻土中的氮素转化,当 C /N 均为 12∶ 1时,反硝化在水稻-小麦轮作土壤中占主导地位,而每年仅种单季稻的土壤中占主导地位的是 DNRA[49]。植物根系释放的 O2 会增强稻田 DNRA,但对反硝化作用没有显著影响,在高 C /N 下,DNRA 对 O2 的敏感性不如反硝化作用[61],与 Rubol 等[62]调查结果一致,DNRA 可能不需要高度还原的环境,但这与 DNRA 需要严格控制大量土壤条件的假说产生矛盾,故须进一步研究来加以验证。现有 DNRA 的研究多集中在海洋水生和河口环境[48]。对于喜铵性作物水稻的氮管理,要维持水稻系统的氮平衡,最大限度地提高氮的利用率,研究水稻土中的 DNRA 过程非常必要。
3 总结与展望
从秸秆还田对稻田微生物群落及功能基因影响的相关研究文献看,国内外学者对水稻土中氮素的转移转化研究主要集中在形态变化,以及土壤理化性质变化对其造成的影响方面,缺乏在关键微生物和功能基因水平上驱动机制的信息,因此,氮素的利用效率和有效转化机制往往被削弱。对于水稻种植过程中何种环境下,哪些功能微生物对氮素的有效转换起主导作用尚不明确。目前,秸秆还田对土壤生态过程的影响研究仍然不够全面系统,笔者认为以下几个方面需要加以更多关注。
1) 土壤微生物与稻田氮素损失的相互关系。土壤的化学和物理性质,如氮源、碳源、水分和 pH 值等的变化会对氮素转化产生影响,但这些性质变化只是通过改变微生物生存环境间接地影响相关微生物的功能,而氮素转化的本质是土壤微生物参与的生物化学过程。因此,研究微生物对氮素转化的作用机理才是研究氮素损失的关键。
2) 秸秆还田下稻田氮素转化功能微生物及功能基因的变化与调控。稻田土壤中氮素转化功能微生物活性与氮素转化基因组成、数量的关系目前并不清楚,尤其是秸秆还田后对氮素转化功能微生物的影响,以及这些微生物的变化是如何来调控氮素转化速率和酶的生成也都不得而知。
3) 氮循环功能微生物的多样性研究。随着分子生物学及相关技术的快速发展,对于氮循环功能微生物的研究已经不仅仅局限于分离培养,而应更多地把目光集中在未分离培养的微生物多样性研究上。总之,深入研究稻田微生物介导的氮素转化,厘清起主导作用的功能微生物、功能基因的变化和驱动因子之间的关系,有利于稻田氮肥减施增效管理,提高稻田生态健康水平。
参考文献( References) :
[1] 杨丽,刘文,兰韬,等. 我国秸秆还田技术与标准的现状研究[J]. 中国农业信息,2017 ( 21) : 12 - 17. YANG L,LIU W,LAN T,et al. Research on the status quo of technology and standard of straw returning in China[J]. China Agricultural Information,2017 ( 21 ) : 12 - 17. ( in Chinese)
[2] 胡乃娟,韩新忠,杨敏芳,等. 秸秆还田对稻麦轮作农田活性有机碳组分含量、酶活性及产量的短期效应[J]. 植物营养与肥料学报,2015,21 ( 2) : 371 - 377. HU N J,HAN X Z,YANG M F,et al. Short-term influence of straw return on the contents of soil organic carbon fractions, enzyme activities and crop yields in rice-wheat rotation farmland[J]. Journal of Plant Nutrition and Fertilizer,2015,21 ( 2) : 371 - 377. ( in Chinese with English abstract)
[3] 董印丽,李振峰,王若伦,等. 华北地区小麦、玉米两季秸秆还田存在问题及对策研究[J]. 中国土壤与肥料,2018 ( 1) : 159 - 163. DONG Y L,LI Z F,WANG R L,et al. Study on the problems and countermeasures of returning wheat and corn stalks into the North China[J]. Soil and Fertilizer Sciences in China, 2018 ( 1) : 159 - 163. ( in Chinese with English abstract)
[4] 何文寿,李生秀,李辉桃. 水稻对铵态氮和硝态氮吸收特性的研究[J]. 中国水稻科学,1998,12 ( 4) : 249 - 252. HE W S,LI S X,LI H T. Characteristics of ammonium and nitrate uptake in rice[J]. Chinese Journal of Rice Science, 1998,12 ( 4 ) : 249 - 252. ( in Chinese with English abstract)
秸秆还田的论文范文你还可以浏览:《玉米秸秆还田技术支付与受偿意愿差异性研究》
转载请注明来自:http://www.lunwenhr.com/hrlwfw/hrnylw/11278.html